Rapp. Comm. int. Mer Médit., 37,2004
150
SIMULATIONS OF THE PHYSICAL AND BIOLOGICAL VARIABILITY AT A NORTHERN ADRIATIC SEA
STATION: THE IMPACT OF TURBULENCE CLOSURE SCHEMES AND BOUNDARY FORCING
Marcello Vichi
1
, Sandro Carniel
2
*, Lakshmi Kantha
3
, Mauro Sclavo
2
1
Istituto Nazionale di Geofisica e Vulcanologia, Via D. Creti, 12, Bologna I-40100, Italy - vichi@bo.ingv.it
2
National Research Council, Istituto per le Scienze del Mare San Polo 1364, Venice I-30125, Italy
* Sandro.Carniel@ve.ismar.cnr.it - Mauro.Sclavo@ve.ismar.cnr.it
3
Department of Aerospace Engineering Sciences, CB 431, University of Colorado, Boulder, CO 80309, United States
kantha@colorado.edu
Abstract
A 1-D model, coupling the physical model GOTM and the ecosystem model ERSEM, has been used to hindcast physical and biological
variability in the northern Adriatic Sea. We studied the impact of the turbulence closure model (TCM) on the thermal structure and
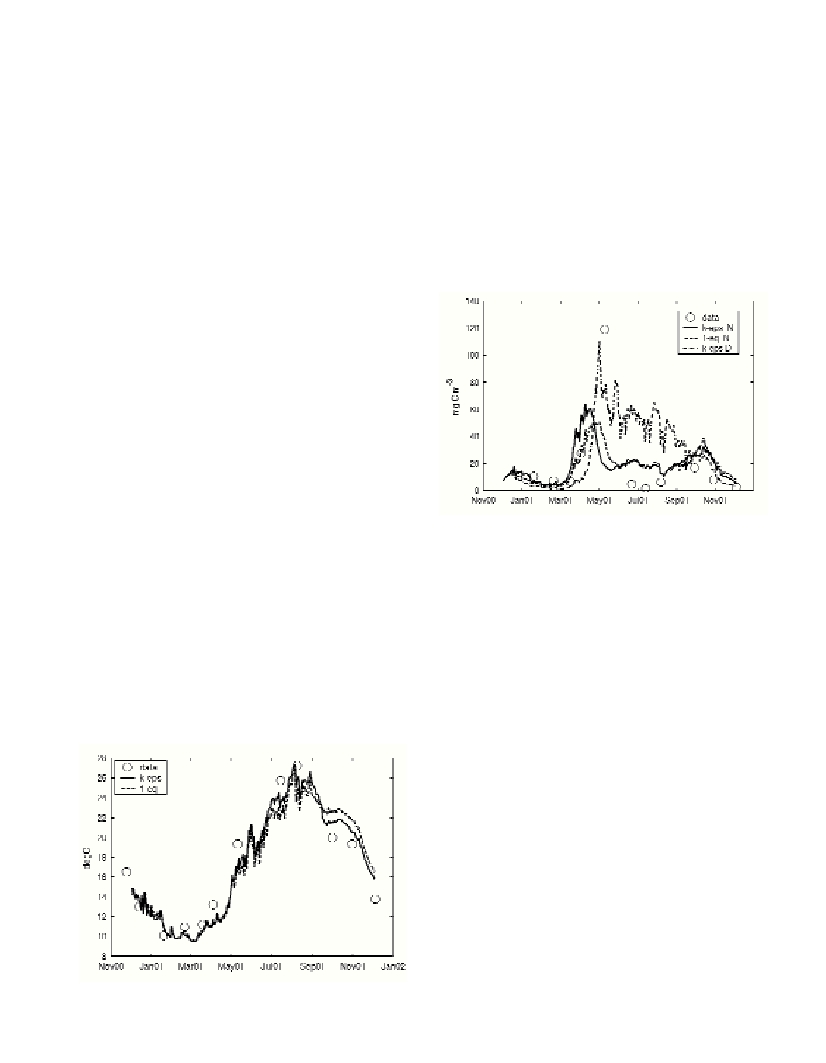
biological productivity. Results compare favorably with physical observations (Fig. 1). The hindcast of biological variables is however less
satisfactory. The results are not sensitive to the second moment TCM used. Changes are however, striking when different boundary
conditions are applied on nutrients (Fig. 2). This suggests that sources of uncertainties other than the TCM, need to be explored to hindcast
the biological state better.
There is a generalized, widely-held view that an accurate
simulation of the physical state is a pre-requisite to the proper
simulation of the biological state. Biology is expected to directly
respond to physical variability, and ecosystem models should be able
to encompass this dynamical behavior. The aim of this work was to
investigate how relevant the description of physical processes is in the
dynamics of shallow ecosystems with respect to other potential
sources of uncertainties.
Therefore, a 1-D coupled physical-biological model has been used
to hindcast the observed seasonal variability for the year 2001 at a
shallow water station in the northern Adriatic Sea of approximately
29m depth (Lat 45.25°N, Long 12.76°E).
The model couples two existing state-of-the-art submodels: the
General Ocean Turbulence Model (GOTM, [1]) and the most recent
version of the European Regional Sea Ecosystem Model (ERSEM III,
[2]).
A relatively high abundance of biological observations are available
at this site for comparison with the model results.
We tested a variety of turbulence closure models (TCMs) and
boundary forcings, and assessed their impact on the simulation of the
thermal structure (Fig.1) and primary producers (e.g. ?agellates,
Fig.2) at the station. The chosen schemes were: 1) k-epsilon; 2)
generic length scale; 2) k-omega and 3) a simple 1-equation model.
Significant differences in the thermal cycle between the model and
the observations become evident only when low-order TCMs are
adopted (see differences between k-epsilon and 1-equation in Fig. 1)
or when the surface forcing is incomplete (e.g., salinity prescribed
only at surface) or when the frequency of surface forcing is changed
(e.g., wind stress from hourly to 6-hourly). Significant differences
also show up in the biochemical variables under these conditions
(Fig.2).
The use of high-frequency physical forcing lets the model achieve
a satisfactory agreement between modeled and measured
temperatures in the water column The results differ only slightly when
different state-of-the-art second order TCMs are used, which suggets
that these TCMs have converged in recent years. Biological variables
in these cases are mostly unaffected by the changes in TCM.
Our conclusions are that, at least in this case, the current status of
physical model is such that it is able to provide a sufficiently accurate
description of the physical state of the water column.
However, the description of the observed biochemical variability is
still far from satisfactory. Therefore, we investigated the importance
of other potential sources of unertainty, as for example the ways of
including the nutrient data as boundary conditions (BC), which are
extremely important in this area due to the presence of the Po river.
Fig. 2 shows how phytoplankton results are strongly sensitive to the
change of BC from von Neumann to Dirichlet methods. Similar
results are obtained if other important model parameters are varied,
such as the ones involved in the C:Chl ratio dynamics.
It appears thus necessary to focus on the refinement of the
parameterization of biochemical processes, and explore the
interactions with other unknown factors such as the proper
assimilation of external inputs.
References
1-Burchard, H., Bolding, K., Villarreal, M.R., 1999. GOTM, a general
ocean turbulence model. Theory, implementation and test cases, European
Commission, Report EUR 18745, 103 p.
2-Vichi, M. Baretta, J.W., Baretta-Bekker, J.G., Ebenhöh, W., Kohlmeier,
C., Ruardij, P. European Regional Seas Ecosystem Model III. Review of
the biogeochemical equations. http://www.bo.ingv.it/ersem3
Fig. 1. Model-data comparison for temperature. Model results obtained
with the k-epsilon and the 1-equation TCMs.
Fig. 2. Model-data comparison for phytoplankton (e.g. ?agellates). k-eps
N = k-epsilon TCM with Neumann BC; 1-eq N = 1-equation TCM with
Neumann BC; k-eps D = k-epsilon TCM with Dirichlet BC