ADHESION OF DIFFERENT MARINE BACTERIA, OUTERMEMBRANE PROTEINS
AND LIPOPOLYSACCHARIDE PRODUCTION
Maria Blažina
1*
, Paolo Landini
2
, Mirjana Najdek
1
, Dragica Fuks
1
and Vesna Svetlicic
3
1
Center for Marine Research, Ruder Boškovic Institute, Rovinj, Croatia,
2
Swiss Federal Institute of Environmental Technology (EAWAG), 8 600 Dübendorf, Switzerland
3
Center for Marine and Environmental Research, Ruder Boškovic Institute, Zagreb, Croatia
* maria.madzarac@cim.irb.hr
Abstract
Marine bacteria were identified by fatty acid analysis and their ability to attach to surfaces was tested. We found that bacteria belonging
to Cytophaga-Flavobacteria(CF) displayed stronger adhesion properties than the
?
-Proteobacteriaisolates, thus suggesting that CFmight
be favoured in aggregate colonisation. These differences are also re?ected in different outer membrane proteins and lipopolysaccharide
composition.
Key words: marine bacteria, adhesion, mucilaginous aggregates, northern Adriatic
Rapp. Comm. int. Mer Médit., 37,2004
265
Introduction
Microbial exopolysaccharides (EPS) are carbohydrate-enriched
polymers produced by microalge and bacteria that bind aggregates
and form dense biofilms near the sediment-water interface. EPS
synthesized by attached bacteria strengthen their binding to surfaces,
but outer membrane proteins (OMP) are required for the early stages
of adhesion (1).
Our aim was to determine the differences in EPS at the cell-surface
of diverse strains commonly found in sediment and mucilaginous
aggregates, and in attachment of these strains to solid surfaces.
Materials and Methods
Six Gram-negative bacterial strains isolated from northern Adriatic
seawater were grown in Marine Broth, at 25°C with vigorous shaking.
Each isolate was saponified, methylated and their fatty-acid (FA)
patterns were analysed by GC/MSD. OMP and LPS of the strains
were extracted and analysed by one-dimensional sodium dodecyl
sulfate-polyacrylamide (SDS-PAGE) and polyacrylamide (PAGE) gel
electrophoresis, respectively (2). For adhesion assays, cultures were
washed and re-suspended in phosphate-buffered saline (PBS). The
suspensions were loaded onto columns filled with pure sea sand
(Fluka), fractions were collected and cell adhesion to sand was
measured as the ratio of the optical density (OD
280
) in each column
fraction to the OD
280
of the initial bacterial suspension (2).
Results and discussion
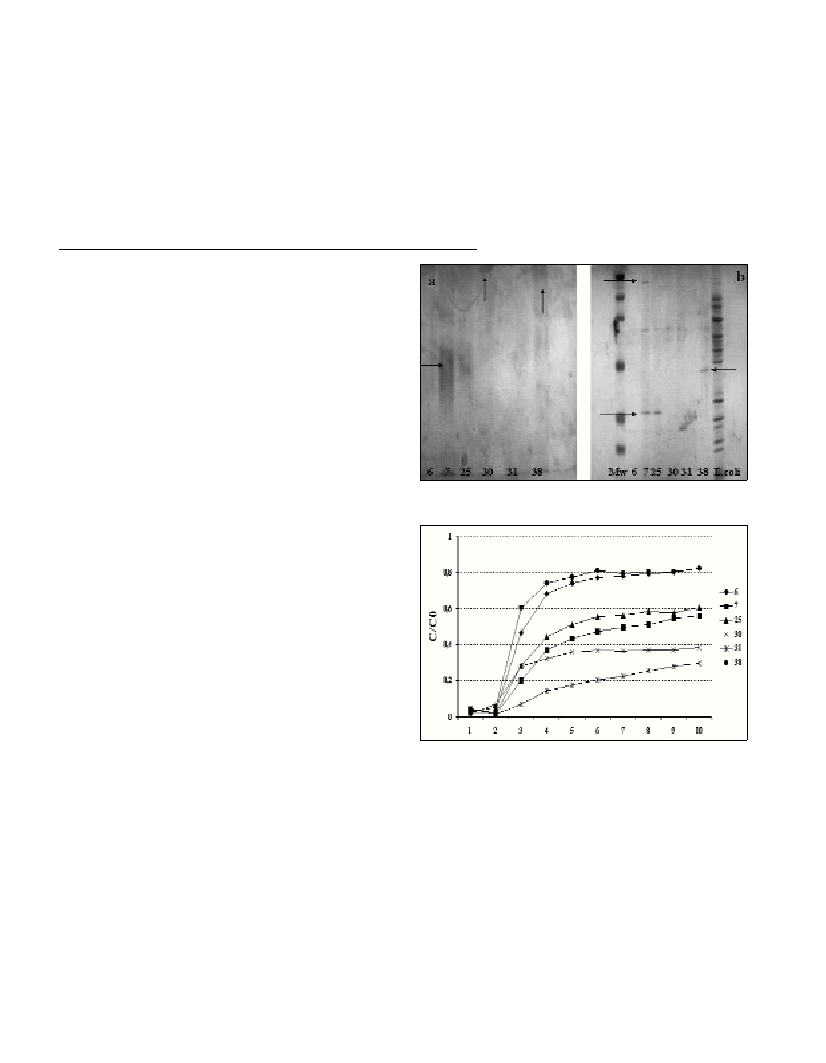
Bacterial isolates had FA patterns corresponding to Cytophaga-
Flavobacteria (7, 25, 30, 31) and Proteobacteria(6, 38). PAGE
analysis of EPS demonstrated a marked difference between marine
bacteria 7/25, 38 and other three strains (Fig. 1). LPS analysis of strain
7 and 25 gave the same band, but slightly more expressed in strain 7.
Strains 30 and 38 had a strong LPS and OMP analysis highlighted the
differences in the extracellular composition of different bacteria:
strains 7 and 25 produced acidic EPS that ran as a smear in PAGE
(Figure 1A). In contrast, strains 30 and 38 produced high MW LPS,
detectable as a dense band at the top of the gel. In addition, strain 38
produced lower MW EPS. Analysis of OMP highlighted the
production of different dominant bands, expressed in strains 7, 25
(around 30 kDa) and 38 (around 40 kDa). In addition, higher MW
bands were present in strains 7 and 25. Differences in the extracellular
structures were re?ected in adhesion properties: adhesion tests
showed efficient attachment by strains 7, 25, 30 and 31 belonging to
the CFgroup while 6 and 38 (Proteobacteria) displayed poor
adherence (Fig. 2).
Favourable adhesive capabilities of Cytophaga-Flavobacteria
indicate that these bacteria are well adapted for colonising solid
surfaces. Since they mainly utilise decomposing refractory macro-
molecules, they can meet their nutritional requirements in the mucous
matrix of aggregates or on epipelic biofilms (3), without having to
compete with Proteobacteria, which mainly utilise lower MW
substances. In mucilaginous aggregates found during periods of un-
usually massive aggregation in northern Adriatic, increasing propor-
tions of branched fatty acids on aged mucilaginous aggregates (4) indi-
cate more efficient aggregate colonization by Cytophaga-Flavobacteria
over other bacterial populations. Differences between CFstrains in
OMP and LPS expression could allow us to define different mecha-
nisms of adhesion control and should be further investigated.
Acknowledgement
The support of the project “Identification of bacterial structures
important for bacterial adhesion using electrochemical adhesion
sensors” (7KRPJO65706) is acknowledged.
Fig. 1. a) PAGE analysis of bacterial LPS, and b) SDS-PAGE analysis of
bacterial OMP.
Fig. 2. Adhesion to a sand column. The C/C
0
value was calculated as the
ratio of bacteria recovered from the ?ow-through column to bacteria
loaded onto the column for each fraction.
References
1-Jucker, B., Harms, H. and Zehnder, A. 1996. Adhesion of the Positively
Charged Bacterium Stenotrophomonas (Xantomonas) maltophilia70401
to Glass and Te?on. J. Bacteriol., 178 (18): 5472-5479.
2-Landini, P. and Zehnder, A. 2002. The Global regulatory hnsGene
Negatively Affects Adhesion to Solid Surfaces by Anaerobically Grown
Escherichia coliby Modulating expression of Flagellar Genes and
Lipopolysaccharide Production. J. Bacteriol., 184 (6): 1522-1529.
3-Kirchman, D.L. 2001. The ecology of Cytophaga-Flavobacteriain
aquatic environments. FEMS Microbiol. Ecol., 39: 91-100.
4-Najdek, M., Degobbis, D., Miokovic, D. and Ivancic, I. 2002. Fatty
acid and phytoplankton compositions of different types of mucilaginous
aggregates in the northern Adriatic. J. Plankton Res., 24 (5): 429-441.