Rapp. Comm. int. Mer Médit., 37,2004
285
RESOURCE AVAILABILITY AND THE GROWTH OF DIFFERENT BACTERIOPLANKTON
Jarone Pinhassi*, Laura Alonso and Josep M. Gasol
Institut de Cičncies del Mar-CMIMA (CSIC), Barcelona, Catalunya, Spain - * jarone@cmima.csic.es
Abstract
The simultaneous use of nutrient limitation and dissolved organic carbon utilization bioassays during different seasons provided insights
into the role of variable supply of resources for the growth of marine bacterioplankton. Furthermore, phylogenetic analyses of bacteria
responding to different treatments in bioassays yield important novel clues to the adaptation of different bacterial groups and species to
particular levels of nutrient availability in the sea. These findings suggest that variable patterns of resource availability and utilization may
aid in explaining the abundance and activity of specific marine bacteria.
Key words: Marine bacterioplankton, Dissolved organic carbon, Nutrient limitation
Introduction
Present inventories of the composition of bacterioplankton offer a
fairly good picture of the overall diversity among bacteria in the sea.
Nevertheless, almost no information exists to explain precisely why
particular groups or species of bacteria are successful. It is reasonable
to assume that several physical, chemical and biological factors act
selectively on the bacterioplankton. Considering the plethora of
possible combinations between these factors it is intriguing that
patterns of preference for particular conditions of some bacteria are
emerging. For example, distinct marine bacterial isolates differ
substantially in their enzymatic capacities, suggesting they are
adapted to utilize different portions of organic matter [1]. In
mesocosm experiments, a few Cytophagaspecies had a high growth
capacity in the presence of elevated protein concentrations [2]. Using
microautoradiography, it was confirmed that marine Cytophagahave
a preference for complex macromolecules, such as protein [3]. To
contribute understanding of how bacterioplankton are affected by
resource availability we report results from a study of the growth of
bacterioplankton in nutrient limitation and dissolved organic carbon
utilization bioassays.
Material and methods
Surface seawater from the NW Mediterranean Sea was collected
monthly from 28 January to 16 September 2003 at a coastal site 40 km
N of Barcelona, Spain (41°40’N, 2°48’E).
The effect of nutrient addition on the growth of heterotrophic
bacteria was examined in samples of 250 ml of unfiltered seawater.
Nutrients were added to final concentrations of 20 µM C (as glucose
or dimethylsulfoniopropionate, DMSP), 2 µM N (NH
4
Cl), and 0.6 µM
P (NaH
2
PO
4
), singly and in all different combinations in duplicates.
Control bottles received no nutrients. After incubation for 24 h at in
situ temperature in the dark, samples for bacterial production,
bacterial abundance and microbial community DNA were collected.
We determined the growth of bacteria combined with
measurements of dissolved organic carbon (DOC) consumption in
duplicate seawater dilution cultures enriched with ammonium and
phosphate as well as in duplicate control cultures without additions.
The bacterial assemblage (0.8 µm-pore-size filtered) was inoculated
into 0.2 µm-pore-size filtered water used as growth medium. Bacterial
growth was monitored until stationary phase (4 d), and subsamples for
bacterial abundance and thymidine and leucine incorporation were
collected twice daily. Microbial community DNA for phylogenetic
analyses was taken at the endpoint. These analyses included PCR-
denaturing gradient gel electrophoresis (DGGE) of 16S rDNA and
sequencing of bands.
Results and discussion
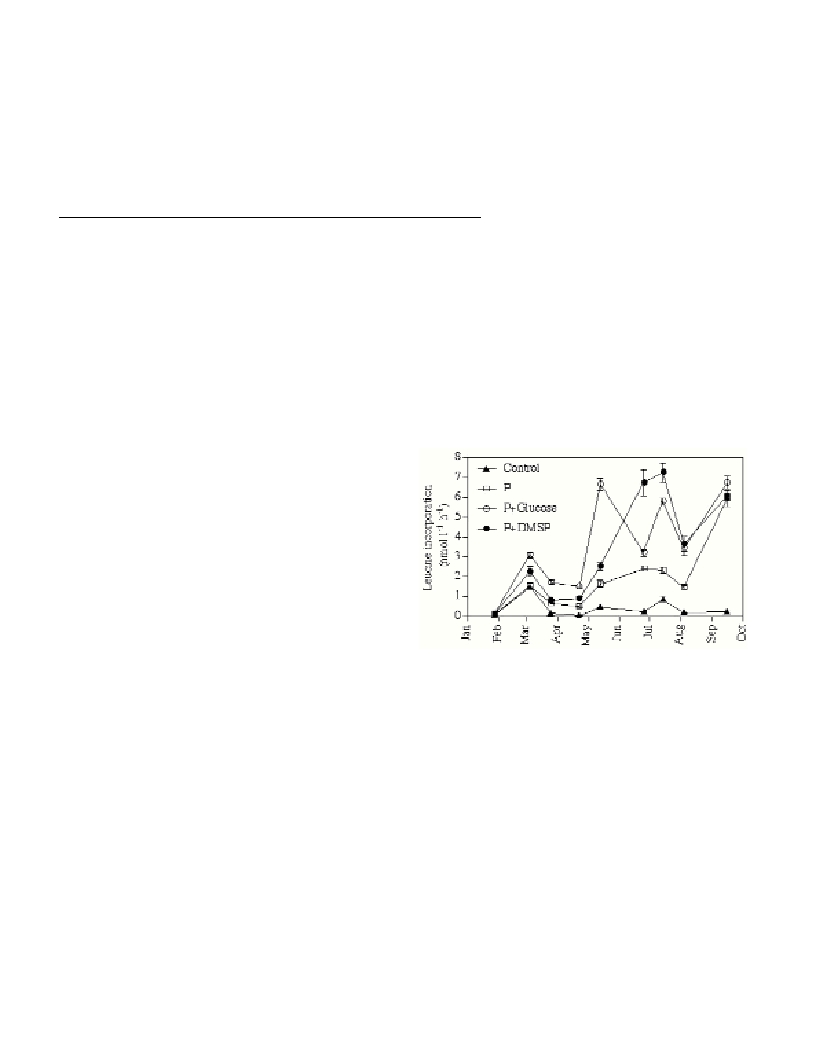
The nutrient addition experiments with unfiltered seawater showed
that bacterioplankton growth was limited by the availability of P
during most of the year (Fig. 1). Notable exceptions occurred during
winter, when rain storms provoked temporary upwelling of deep water
and/or land runoff that provided bacteria with nutrients. Combined
addition of C, either as glucose or DMSP, and P consistently yielded
higher growth compared to only P (Fig. 1). This suggests that
although labile DOC accumulates in this sea area during summer (see
[4] and below) it is less easily utilized compared to glucose or DMSP.
Our data also indicate that DMSP is a highly preferred source of
organic carbon to marine bacteria.
Bacteria in the control dilution cultures ranged between 0.7 to
1.2 x 10
6
cells ml
-1
, with highest values in winter. During winter,
growth of bacteria in the enriched dilution cultures was only slightly
higher than in the controls, while in spring and summer enrichment
caused up to fourfold higher bacterial growth rates and abundance.
These data corroborate that DOC accumulates in surface seawater in
regions where bacterioplankton growth is limited by mineral
nutrients[4].
Analyses of bacterioplankton composition in our bioassays showed
that some bacteria abundant in the initial water also grew in the
control dilution cultures - particularly Roseobacterand Cytophaga
relatives. Although these groups are frequently detected as abundant
in seawater using DGGE and FISH, it was found that specific
members of these groups became dominant in the cultures depending
on nutrient status and season. In the enriched cultures, fast-growing
gamma-proteobacteria, such as Alteromonas, quickly became
dominant. Overall, there was a intimate relationship between the
amount of DOC consumed and the composition of the bacterial
assemblage.
Fig. 1. Temporal changes of the response in bacterial production to the
addition of nutrients. Treatments without P behaved as the control.
Acknowledgements
We thank Laura Gómez Consarnau and Elena Blanc for skilful
technical assistance. This work was supported by the EU project
BASICS grant EVK3-CT-2002-00078.
References
1-Martinez, J., Smith, D.C., Steward, G.F. and Azam, F., 1996.
Variability in ectohydrolytic enzyme activities of pelagic marine bacteria
and its significance for substrate processing in the sea. Aquat. Microb.
Ecol.,10: 223-230.
2-Pinhassi, J., Azam, F., Hemphälä, J., Long, R.A., Martinez, J., Zweifel,
U.L. and Hagström, Ĺ., 1999. Coupling between bacterioplankton species
composition, population dynamics, and organic matter degradation.
Aquat. Microb. Ecol.,17: 13-26.
3-Cottrell, M.T. and Kirchman, D.L., 2000. Natural assemblages of
marine Proteobacteria and members of the Cytophaga-Flavobactercluster
consuming low- and high-molecular-weight dissolved organic matter.
Appl. Environ. Microbiol., 66: 1692-1697.
4-Thingstad, T.F., Hagström, Ĺ. and Rassoulzadegan, F., 1997.
Accumulation of degradable DOC in surface waters: Is it caused by a
malfunctioning microbial loop? Limnol.Oceanogr., 42: 398-404.