ANTIOXIDANT DEFENCES IN COASTAL AND DEEP-SEA FISH: A COMPARATIVE STUDY
Ramón Lavado *, Luz M. García de la Parra, Estefanía Escartín, Cinta Porte
Environmental Chemistry Dept. IIQAB-CSIC, Barcelona, Spain - * rlpqam@cid.csic.es
Abstract
Rather low levels of oxidative stress are expected in deep-sea organisms due to both, their reduced metabolic rate and the physicochemical
conditions of a dark, poorly oxygenated environment However, when antioxidant defences of coastal and deep-sea species collected from
the NW Mediterranean were compared, only the activity of glutathione peroxidase was observed to decrease with depth; catalase and
superoxide dismutate remained unchanged. Thus, dangers associated to reactive oxygen species (ROS) exposure did not appear to decrease
in deep-sea areas, and other factors (presence of swim bladder, diet, pollutant exposure) can significantly enhance the endogenous
production of ROS in deep-sea organisms.
Keywords: deep-sea fish, catalase, superoxide dismutase, glutathione peroxidase
Rapp. Comm. int. Mer Médit., 37,2004
385
Several reports suggest that marine organisms are exposed to high
environmental concentrations of potentially deleterious oxygen
derivatives, viz. superoxide anion, hydrogen peroxide. These reactive
oxygen species (ROS) are abundant in the upper layers of the oceans,
but their concentrations decreases with increasing depth. In deeper
regions, the exposure to oxidative stress is considerable lower because
of reduced light irradiance, lower oxygen levels; and reduced
metabolic activity. ROS can oxidize most cellular constituents, such
as DNA, proteins, and lipids, and markedly affect the physiology of
the cell, leading to the initiation of cancer and cellular death (1).
Consequently, organisms have developed defence systems against
oxidative damage, consisting of antioxidant scavengers (glutathione,
vitamin C, vitamin E, carotenoid pigments), and specific antioxidant
enzymes: catalase (CAT), superoxide dismutase (SOD), and
glutathione peroxidase (GPX).
This work was designed as a comparative study on the presence of
antioxidant enzymes in coastal (30-50 m) and deep-sea fish (1500-
1800 m depth) from the NW Mediterranean. Special attention was
paid to the distribution of antioxidant enzyme systems in deep-sea fish
species, and whether the reduced danger linked to oxygen toxicity
could have lead to a reduction of their biochemical defences against
oxidative damage. To test this hypothesis we investigated the activity
of the antioxidant enzymes (CAT, SOD, GPX) in the liver of 10 fish
species selected on the basis of their abundance, commercial interest
and habitat. Mullus barbatus, Serranus cabrilla, Serranus hepatus,
Sparus aurata, Diplodus annularis, Scorpaena porcus, and Solea
vulgariswere collected at 30-50 m depth; Lepidion lepidion,
Coryphaenoides guentheriand Bathypterois mediterraneusat 1500-
1800 m. The number of individuals analyzed varied between 10 and
70, depending on the species. The methods used for the determination
of antioxidant enzymes are described in Porte et al.(2).
No significant differences (Anova test, Tukey-Karamer multiple
comparisons test, P>0.05) between coastal and deep-sea species were
observed in terms of SOD activity; the highest activity detected in S.
hepatus (18.4 ±2.2 units/mg protein), and the lowest in S. aurata(5.4
±0.6 units/mg protein). Intermediate values were recorded for deep-
sea species (8-14 units/mg protein). SOD converts O
2.–
into H
2
O
2
,
which can in turn be detoxified into water and oxygen by either CAT
or GPX, which utilizes glutathione as an electron donor. There was a
significant correlation (r=0.85; P<0.05) between the activities of SOD
and CAT in the studied species. CAT, which detoxifies H
2
O
2
into
water and oxygen, was determined in liver cytosol (broken
peroxisomes) or in peroxisomes + cytosol (total activity), but no
relationship with depth was observed. In agreement with other studies
(3), the most active species, from both coastal (S. aurata) and deep-
sea areas (L. lepidion, C. guentheri), showed higher CAT activities
than the less active ones, with reduced motility (S. cabrilla, S. porcus,
B. mediterraneus).
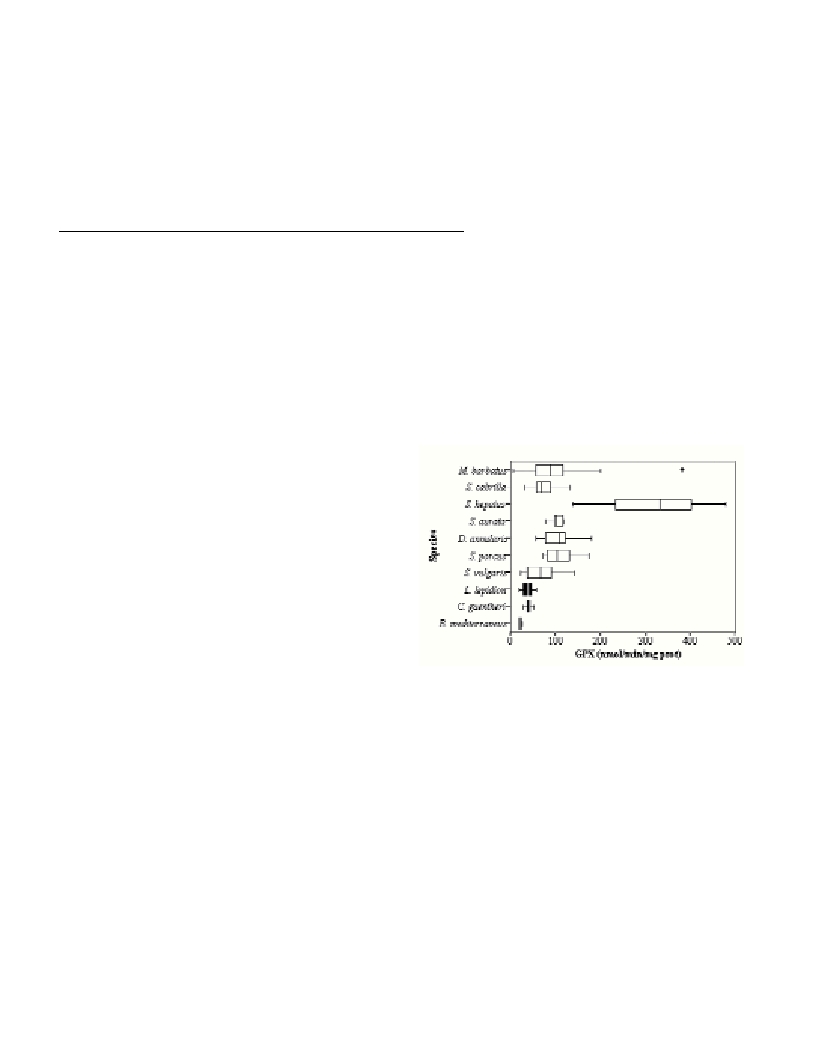
In contrast, the activity of GPX using H
2
O
2
as a substrate decreased
with depth. In shallow species, GPX ranged from 74 to 110
nmol/min/mg protein, with the exception of S. hepatus, that had an
activity of 340 nmol/min/mg protein. Lower activities were recorded
in deep-sea specimens (22-40 nmol/min/mg protein) (Fig. 1). The
reason for this apparent preference for CAT in deep-sea species might
be related to the limited resources available in the deep-sea, and to the
fact that CAT requires neither cofactors nor energy to detoxify H
2
O
2
,
while GPX consumes glutathione, which is oxidized and must then be
recycled by a NADPH-consuming enzyme (glutathione reductase).
Despite of the reduced metabolic requirements of deep-sea fish
species, this study shows that the dangers associated to ROS exposure
do not decrease with increasing depth. Other factors, such as habitat,
diet, exposure to pollutants, and the presence of swim bladder, can
significantly increase the endogenous levels of ROS deep-sea
organisms are exposed to (3). Hence, B. mediterraneus, a sedentary
fish, well adapted to the oligotrophic deep environment, exhibited low
activities of both catalase and GPX (both detoxify H
2
O
2
) in
comparison with the other species, and this may be related to the
absence of a swim bladder. It is reported that with increasing depth
and increasing hydrostatic pressure, most species maintain swim
bladder volume constant by increasing mainly its oxygen content that
may make up to 90% of the gas mixture in deep-sea fish. Thus, the gas
gland tissue operates under conditions of hyperoxia, and this enhances
oxyradical production (4).
Fig. 1. Box plot of glutathione peroxidase (GPX) activity determined in
liver cytosol of different fish species sampled in the NW Mediterranean.
Black boxes indicate deep-sea fish species.
References
1-Cadenas, E., 1995. Mechanisms of oxygen activation and reactive
oxygen species detoxification. Pp. 1-61. In: Ahmad S. (ed.), Oxidative-
Induced Stress and Antioxidant Defences in Biological Systems. Chapman
& Hall, NY.
2-Porte C., Escartín E., García L.M., Solé M., Albaigés J., 2000.
Xenobiotic metabolising enzymes and antioxidant defences in deep-sea
fish: Relationship with contaminant body burden. Mar. Ecol. Prog. Ser.,
192: 259-266.
3-Wilhelm Filho D., Giulivi C., and Boveris A., 1993. Antioxidant
defences in marine fish- I. Teleosts. Comp. Biochem. Physiol., 106C: 409-
413.
4-Jones, D.P., 1985. The role of oxygen concentration in oxidative stress:
hypoxic and hyperoxic stress. Pp. 151-195. In: Sies H. (ed.), Oxidative
Stress. Academic Press, NY.