Rapp. Comm. int. Mer Médit., 36,2001
188
Oxygen and sunshine (UV) are two major genotoxic agents that all
photosynthetic organisms are obliged to be exposed to. One way to
assess the importance of enhanced UV irradiation as a potential source
of biological damage in marine organisms is to determine UV- induced
DNA damage and the capacity for DNA repair. Cyclobutane pyrimi-
dine dimers (CPDs) are exclusive UV product and make 75% of the
UV-induced DNA damage products (1), having an inhibitory effect on
transcription and replication as well as mutagenic effect. We measured
incidence of CPDs in marine phytoplankton Isochrysis galbanacul-
tures (2) grown under different light/dark regimes after artificial UV
exposure. CPD content in I. galbanacells was expressed as SSF
(strand scission factor) - measure of CPD sites as single strand breaks
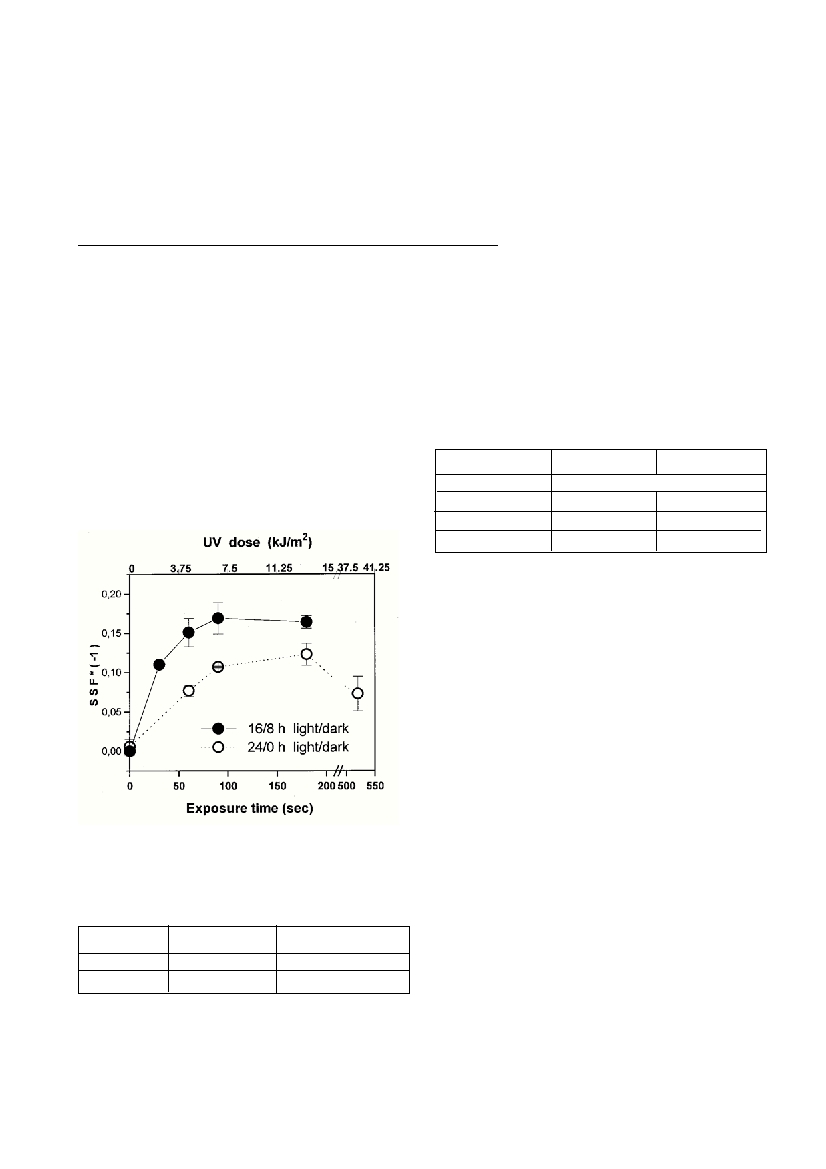
(3) after T4endonuclease V digestion. In cells grown under 16/8 h
(light/dark) regime after exposure to artificial UV (0-37.5 kJ/m
2
) more
CPDs was detected than in the cells grown under constant illumination
(24 h) ( Fig.1).The latter contained higher amount of photosynthetic
pigments (chlorophyll a) as well as UV absorbing compounds (Table
1). These results revealed correlation between light exposure, UV-
absorbing compounds content and sensitivity to UV light, suggesting
strong in?uence of photoprotective compounds in reducing the sus-
ceptibility to UV damage in phytoplankton cells.
Besides protection there is a great importance of the existence of
DNA repair mechanisms as a part of biological response of phyto-
plankton cells to UV irradiation. DNA photodamage and DNA repair
capacity in I. galbanacells exposed to ambient solar irradiation was
measured on August 6 and 20,1996 (Table 2). After 1 hour, DNA dam-
age increased and after 3 hours of exposure, it decreased to the 40% of
the damage obtained after first 1 hour.Thus, DNA protection and dam-
age elimination mechanisms successfully lowered CPDs accumula-
tion, suggesting that sunlight exposure induces DNA damage that
could be repaired even when the DNA damage and repair are happen-
ing simultaneously.
References
1. Mitchell D.L. and Nairn R.S., 1989. The biology of (6-4)
photoproduct.Photochem. Photobiol., 49(6): 805-819
2. Fafandel M, Bihari N, Krajcar V., Müller W.E.G., Zahn R.K., Batel R.,
2001.Specific detection of cyclobutane pyrimidine dimers in
phytoplankton DNA by non-radioactive assay based on T 4endonuclease
V digestion. Sci.Tot. Env.(in press)
3.Fafandel M.., 1997.Effect of UV irradiation on phytoplankton
Isochrysis galbanaParke DNA grown in monoculture. M. Sc. Thesis,
Zagreb, 1997 ( in Croatian)
Rapp. Comm. int. Mer Médit., 36,2001
CYCLOBUTANE PYRIMIDINE DIMERS DETECTION IN MARINE PHYTOPLANKTON
ISOCHRYSIS GALBANAFOLLOWING UV IRRADIATION
Maja Fafandel*, Nevenka Bihari, Renato Batel
Rudjer Boskovic Institute, Center for Marine Research, Rovinj, Croatia - maja@cim.irb.hr
Abstract
The effect of artificial and solar UV irradiation on DNA of marine phytoplankton Isochrysis galbanawas investigated. Cultures grown
under 16/8 light/dark regime contained smaller amount of photosynthetic pigments and UV absorbing compounds and showed greater
sensitivity to UV light than the cultures grown under 24/0 light regime. For specific detection of cyclobutane pyrimidine dimers (CPDs)
inI. galbana alkaline DNA filter elution using T4 endonuclease V was adapted. Artificial UV irradiation increases CPDs in dose-dependant
manner.After 1 h of sunlight exposure, the level of DNA damage increase significantly but prolonged exposure decreased it due to the
efficiency of I. galbanaDNA repair mechanisms.
KEYWORDS: phytoplankton, pigments, UV, sunlight, cyclobutan pyrimidine dimers
Fig.1.DependenceofSStoartificialUVradiationfor
Isochrysis galbanacultures grown under differents light/dark
regimes
Table1. Chlorophyll a and UV-absorbing compounds content in
Isochrysisgalbanacellsgrownunderdifferentlight/dark
IlluminationChlorophyllaUV-absorbingcompo-
n
e
n
t
s
light/dark (h)(µg/10
6
cells)
(A
300
/10
7
cells)
24/0
0.353
0.792
Table 2. CPDs accumulation and their repair in Isochrysis gal-
banacells during sunlight exposure in the Northern Adriatic
(45o 05` N, 13o 30` E, Rovinj, Croatia).
SUNLIGHT EXPOSUREAugust 6, 1996August 20, 1996
Time/h
11:00
-0.006 ± 0,019-0,006 ± 0,019
12:00
-0,055 ± 0,015-0,037± 0,002
14:00
-0,007 ± 0,018-0,015 ± 0,003
SSF